Голги комплекс: опис
Комплекс Голгија откривен је 1898. године. Ова мембранска структура је дизајнирана да излучује једињења која се синтетизују у ендоплазматском ретикулуму. Даље ћемо се упознати са овим системом. 
Комплекс Голги: зграда
Уређај је сноп мембранских дискова у облику диска. Ове врећице су донекле проширене према рубовима. Голги систем мехурића је повезан са тенковима. У ћивотињским ћелијама присутна је једна велика или неколико гомила, које су повезане цевима, у биљне ћелије Дицтиосомес се детектују (неколико одвојених стогова). Комплекс Голги обухвата три одељења. Окружени су мембранским мехурићима:
- цис-најближи нуклеусу;
- медиал;
- транце одељење (најдаље од језгра).
Ови системи се одликују ензимским сетом. У цис одељењу, прва кеса се назива "спасилачки резервоар". Уз његову помоћ, рецептори који долазе из ендоплазматске средње мреже, враћају се назад. Одељење ензима цис се зове фосфогликозидаза. Додаје фосфат манози (угљени хидрати). У средњем делу налазе се два ензима. Ово, нарочито, меннадиасис и Н-ацетилглукозамин трансфераза. Ово последње додаје гликозамине. Ензими транс-одељења: пептидаза (спроводи протеолизу) и трансферазе (уз помоћ трансфера хемијских група). 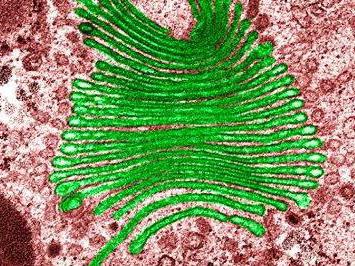
Голги комплекс: функције
Ова структура обезбеђује раздвајање протеина у следећа три тока:
- Лисосомал. Према томе, глицирани протеини продиру у цис-одјел Голги апарата. Неке од њих су фосфорилисане. Као резултат, формира се манозо-6-фосфат - тржишососомски ензими. У будућности, ови фосфатни протеини ће ући у лизосоме и неће бити модификовани.
- Конститутивна егзоцитоза (секреција). Ова струја обухвата протеине и липиде, који су постали компоненте апарата на површини ћелије, укључујући гликокаликс. Такође, могу бити присутна једињења која су део екстрацелуларног матрикса.
- Индуцед сецретион. Протеини, који функционишу изван ћелије, површински апарат, продиру у тај ток у унутрашњем окружењу тела. Индуцирана секреција је карактеристична за секреторне ћелије.
Голгијев комплекс учествује у формирању мукозних секрета - мукополисахарида (гликозаминогликана). Уређај такођер формира угљикохидратне компоненте гликокаликса. Углавном су то гликолипиди. Систем такође обезбеђује сулфатацију протеина и елемената угљених хидрата. Комплекс Голги је укључен у парцијалну протеолизу протеина. У неким случајевима, због тога, једињење из неактивног улази у активну форму (на пример, проинзулин се трансформише у инсулин). 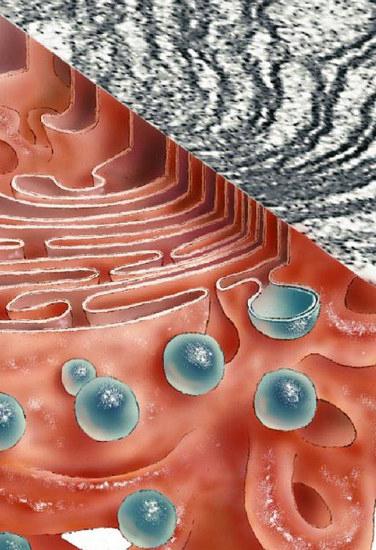
Покретање једињења из ендоплазматског ретикулума (ЕПС)
Комплекс је асиметричан. Налази се ближе језгру ћелије и обухвата најзрелије протеине. Везикуле, мембранске везикуле, стално су причвршћене за ове врећице. Они избијају из ендоплазматског грануларног ретикулума. На његовим мембранама пролази процес синтезе протеина рибозомима. Транспорт једињења из ендоплазматског ретикулума у Голги комплекс је неселективан. Истовремено, нетачно или непотпуно пресавијени протеини остају у ЕПС-у. Обрнуто кретање једињења у ендоплазматски ретикулум захтева посебну сигналну секвенцу и омогућено је везивањем ових супстанци за мембранске рецепторе у цис-компартменту.
Протеин модифицатион
У резервоарима комплекса сазревају се једињења која су намењена за излучивање, трансмембранске, лизосомалне и друге супстанце. Ови протеини се сукцесивно крећу дуж тенкова према органелима. Њихове модификације почињу у њима - фосфолирање и гликозилација. Током првог процеса, остатак је везан за протеине. фосфорна киселина. У О-гликозилацији, комплексни шећери су усидрени преко атома кисеоника. Различити резервоари садрже различите каталитичке ензиме. Сходно томе, сукцесивно се одвијају различити процеси са сазријевањем протеина у њима. Несумњиво, такав степен феномена треба контролисати. Као врста "ознаке квалитета" користе се полисахаридни остаци (углавном маноза). Означавају зреле протеине. Даљње кретање кроз резервоаре једињења није у потпуности схваћено од стране науке, упркос чињеници да отпорне материје остају мање или више повезане са једном врећом. 
Транспорт протеина из апарата
Од мјешовитих мјехурића. Садрже потпуно зрела протеинска једињења. Главна функција комплекса је сортирање протеина који пролазе кроз њега. У апарату, формирање "тространог протеинског тока" - сазревање и транспорт:
- Цомпоундс плазма мембрана.
- Сецретс.
- Лизосомални ензими.
Кроз везикуларни транспорт, протеини који су прошли кроз Голги комплекс се достављају на одређене локације у складу са "ознакама". Овај процес такође није у потпуности схваћен у науци. Утврђено је да транспорт протеина из комплекса захтева учешће специфичних мембранских рецептора. Они препознају једињење и обезбеђују селективно пристајање везикуле и једне или друге органеле. 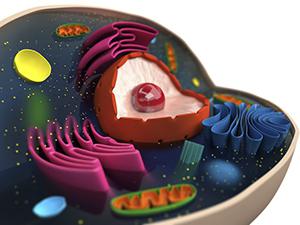
Формирање лизосома
Кроз апарат пролази много хидролизних ензима. Додавање горе наведене етикете врши се уз учешће два ензима. Специфично препознавање лизосомалних хидролаза елементима њихове терцијарне структуре и додавање Н-ацетилглукозамин фосфата врши се Н-ацетилглукозамин фосфотрансферазом. Фосфогликозид, други ензим, уклања Н-ацетилглукозамин, што доводи до формирања ознаке М6Ф. Она се, пак, препознаје од рецептора. Уз његову помоћ, хидролазе се достављају везикулама и њиховим мандатима у лизосоме. У киселим условима, фосфат се одваја од зреле хидролазе у њима. Ако постоје поремећаји у активности Н-ацетилглукозамин фосфотрансферазе услед мутација или због генетских дефеката у М6Ф рецептору, сви лизосомални ензими се достављају према спољашњој мембрани. Затим се излучују у ванћелијске услове. Такође је утврђено да се неки од М6П рецептора такође транспортују до спољашње мембране. Они врше повратак случајно ухваћених лизосомалних ензима из спољашњег окружења унутар ћелије током ендоцитозе. 
Транспорт супстанци до спољашње мембране
Обично, чак иу фази синтезе, протеинска једињења спољашње мембране са њиховим хидрофобним регионима су уграђена у зид ендоплазматског ретикулума. Затим се испоручују у комплекс Голги. Одатле се транспортују до површине ћелије. У процесу фузије плазмалеме и везикула, таква једињења се не ослобађају у околину.
Сецретион
Скоро сва једињења произведена у ћелији (и протеинске и не-протеинске природе) пролазе кроз Голги комплекс. Тамо се развијају у секреторне везикуле. У погонима са дицтиос, материјал се тако производи. ћелијски зид.