Гене Дрифт - Еволутион Фацтор: Разлог, Вриједност, Примјери
Поред природне селекције, постоји још један фактор који може утицати на повећање садржаја мутантног гена. У неким случајевима, може чак и да смањи нормални алел. Овај феномен се назива "померање гена у популацији". Размотримо детаљније шта је то процес и какве су његове посљедице. 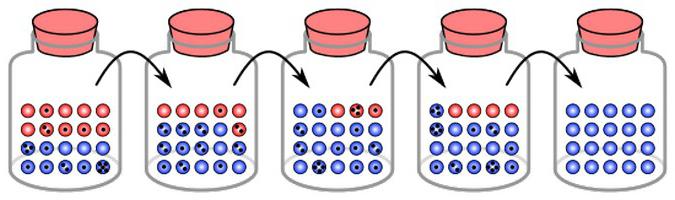
Опште информације
Одвијање гена, примјери који ће бити наведени у чланку испод, су одређене промјене које се биљеже из генерације у генерацију. Сматра се да овај феномен има своје механизме. Неки истраживачи су забринути да у генском фонду многих, ако не и свих нација, количина аномалних гена који се појављују тренутно расте прилично брзо. Они одређују наследну патологију, формирају предуслове за развој многих других болести. Такође се верује да је патоморфоза (промена знакова) разних болести, укључујући и оне менталног карактера, управо заношење гена. Појава о којој се ради се одвија великом брзином. Као резултат, број ментални поремећаји узима непознате облике, постаје непрепознатљив у поређењу са њиховим описом у класичним издањима. Истовремено, значајне промене се директно уочавају у структури саме психијатријске болести. На пример, померање гена брише неке облике шизофреније које су се среле раније. Уместо тога, постоје патологије које се тешко могу одредити модерним класификаторима. 
Вригхтова теорија
Случајни генски помак проучаван је помоћу математичких модела. Користећи овај принцип, Вригхт је извео теорију. Он је сматрао да је пресудна важност померања гена под константним условима уочена у малим групама. Они постају хомозиготни и варијабилност опада. Вригхт је такође веровао да, као резултат промена у групама, могу да се формирају негативне наследне карактеристике. Као резултат тога, цјелокупно становништво може умријети без доприноса развој врсте. Истовремено, селекција игра велику улогу у многим групама. У том смислу, генетске варијације унутар популације ће опет бити безначајне. Постепено, група ће се добро прилагодити условима животне средине. Међутим, накнадне еволутивне промјене ће зависити од појаве повољних мутација. Ови процеси су прилично спори. У том смислу, еволуција великих популација није карактерисана великом брзином. У групама средњих вредности уочава се повећана варијабилност. Истовремено, формирање нових корисних гена се догађа случајно, што заузврат убрзава еволуцију. 
Вригхтови закључци
Када се из популације изгуби један алел, може се појавити због одређене мутације. Али ако је врста подељена у неколико група, у једном од којих недостаје један елемент, у другом, други, онда ген може да мигрира одатле где је, до тамо где није. Дакле, варијабилност ће остати. С обзиром на то, Вригхт је закључио да ће се развој брже одвијати у оним врстама које су подијељене на бројне различите популације. Истовремено, између њих је могућа нека миграција. Вригхт се сложио с тим природна селекција игра веома значајну улогу. Међутим, уз овај резултат еволуције долази и до гена. Он идентификује трајне промене унутар приказа. Поред тога, Вригхт је веровао да су многе карактеристичне особине које су настале због заношења индиферентне, ау неким случајевима чак и штетне за одрживост организама. 
Спорови истраживача
Било је неколико мишљења о Вригхтовој теорији. На пример, Добжански је веровао да је бесмислено постављати питање који је од фактора значајнији - природна селекција или генетски занос. Он је то објаснио својом интеракцијом. У суштини, могуће су следеће ситуације:
Ad
- Ако у развоју једне или друге врсте селекција заузме водећу позицију, или ће се означити смерна промена у фреквенцијама гена или стабилно стање. Потоње ће бити одређено условима животне средине.
- Ако је током дужег периода померање гена значајније, онда усмерене промене неће бити узроковане природним окружењем. Истовремено, неповољни знаци, чак иу малим количинама, могу се у групи довољно проширити.
Треба, међутим, напоменути да сам процес промене, као и узрок заношења гена, данас није довољно истражен. У том смислу, не постоји јединствено и конкретно мишљење о овој појави у науци.
Ad
Одвагање гена је фактор еволуције
Због промена, примећује се промена фреквенција алела. То ће се десити док не достигну стање равнотеже. То јест, заношење гена је изолација једног елемента и фиксација другог. У различитим групама, такве промене се дешавају независно. У том смислу, резултати генетског дрифта у различитим популацијама су различити. На крају, један сет елемената је фиксиран у неким, други у другима. Диференцијација гена, с једне стране, стога доводи до смањења разноликости. Међутим, у исто вријеме, она такођер узрокује разлике између група, разлике на неким основама. Ово, заузврат, може послужити као основа за специјацију. 
Однос утицаја
У процесу развоја, генетски дрифт је у интеракцији са другим факторима. Пре свега, веза се успоставља са природном селекцијом. Однос доприноса ових фактора зависи од више околности. Прво, то је одређено интензитетом селекције. Друга околност је величина групе. Дакле, ако су интензитет и снага високи, случајни процеси имају занемарљив утицај на динамику генетских фреквенција. Истовремено, у малим групама са незнатним разликама у способности, ефекат промена је неупоредиво већи. У таквим случајевима, може се поправити мање адаптивни алел, док ће адаптивни бити изгубљен.
Ad
Ефекти промене
Један од главних резултата генетског дрифта је осиромашење различитости унутар групе. То је због губитка неких алела и фиксације других. Процес мутације, напротив, доприноси обогаћивању генетичке разноликости унутар популација. Због мутације, изгубљени алел се може појавити изнова и изнова. С обзиром на чињеницу да је генетски дрифт смјерни процес, истовремено са смањењем интрапопулацијске разноликости, разлика између локалних група се повећава. Противљење овом феномену је миграција. Дакле, ако је у једној популацији алел "А" фиксиран, ау другом - "а", онда се унутар ових група различитост поново појављује. 
Коначни резултат
Резултат генетског дрифта биће потпуна елиминација једног алела и консолидација другог. Што се елемент чешће појављује у групи, већа је вјероватноћа његове фиксације. Као што неки прорачуни показују, могућност везивања једнака је учесталости алела у популацији.
Ad
Мутације
Они се јављају у просеку са учесталошћу од 10-5 по гену по гамету по генерацији. Сви алели који се налазе у групама су се једном појавили услед мутација. Што је популација мања, то је мања вјероватноћа да ће свака генерација имати барем једну особу - носиоца нове мутације. Са бројем сто хиљада, свака група потомака са вероватноћом блиском јединству има мутирани алел. Међутим, њена учесталост у популацији, као и могућност њеног фиксирања ће бити прилично ниска. Вјероватноћа да ће се иста мутација појавити у истој генерацији, барем код једне особе у популацији од 10, је занемарљива. Међутим, ако се догоди у овој популацији, учесталост мутантног алела (1 у 20 алела), као и шансе за његово фиксирање, биће релативно висока. Код великих популација појављивање новог елемента се дешава релативно брзо. У исто време, његова фиксација је спора. Насупрот томе, мале популације очекују дугу мутацију. Али након његовог појављивања, причвршћивање брзо пролази. Из овога се може извести следећи закључак: шанса за фиксирање неутралних алела зависи само од учесталости мутације. Истовремено, величина популације не утиче на овај процес.
Ad
Молецулар Цлоцк
Због чињенице да је учесталост појављивања неутралних мутација у различитим врстама приближно једнака, брзина фиксирања би такође требала бити приближно једнака. Из овога следи да број промена акумулираних у једном гену треба да буде у корелацији са временом независне еволуције ових врста. Другим речима, што дуже буде период од тренутка раздвајања две врсте са једног предака, то ће више разликовати мутацијске супституције. Овај принцип лежи у основи молекуларног еволуционог сата. Ово дефинише време које је прошло од тренутка када су се претходне генерације разних систематских група почеле развијати независно, независно једна од друге.
Поллинг и Цукуркенд студија
Ова два америчка научника су открили да је број разлика у секвенци аминокиселина у цитокрому и хемоглобину у одређеним врстама сисаваца виши, раније су се њихови еволутивни путеви дивергирали. Након тога, овај узорак је потврђен великом количином експерименталних података. Материјал је укључивао на десетине различитих гена и неколико стотина врста животиња, микроорганизама и биљака. Показало се да се ток молекуларних сати изводи константном брзином. Ово откриће, у ствари, потврђује теорија о којој се ради. Сат је калибриран одвојено за сваки ген. То је због чињенице да је учесталост појаве неутралних мутација у њима различита. У ту сврху се процењује број супституција акумулираних у одређеном гену у таксономима. Њихово време дивергенције се поуздано утврђује коришћењем палеонтолошких података. Након што је калибрисан молекуларни сат, они се могу даље користити. Посебно, уз њихову помоћ, лако је измјерити вријеме у којем је дошло до дивергенције (дивергенције) између различитих таксона. Ово је могуће чак и ако њихов заједнички предак још није идентификован у фосилним записима.