Механизам контракције мишића кратко
Процеси мишићног рада су вишеслојни комплекс физиолошких и биохемијских функција које су од виталног значаја за потпуно функционисање људског тела. Екстерно, такви процеси се могу посматрати у примјерима добровољних покрета при ходању, трчању, мијењању израза лица итд. Међутим, они покривају много већи спектар функција, укључујући и рад респираторног апарата, органа за варење и система за излучивање. У сваком случају, механизам контракције мишића је подржан радом милиона ћелија, у које су укључени хемијски елементи и физичка влакна.
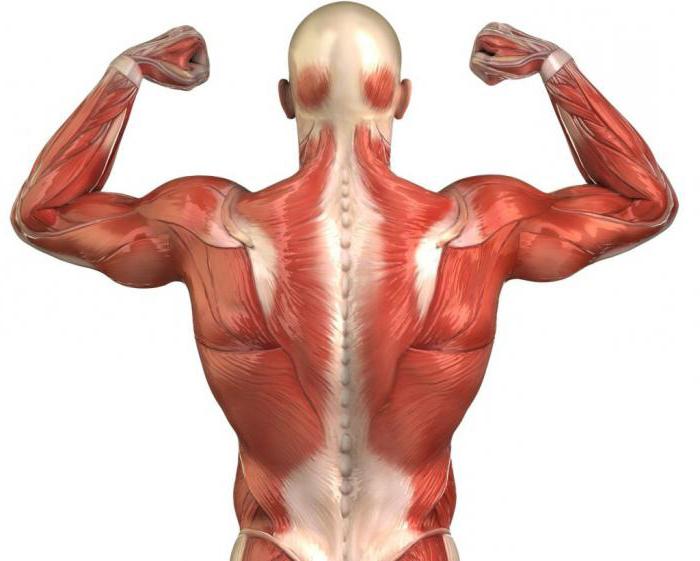
Структурна организација мишића
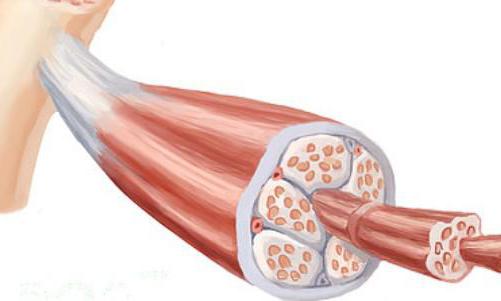
Мишићи се формирају од различитих ткивних влакана која имају тачке везивања за кости скелета. Налазе се паралелно и међусобно комуницирају у процесу мишићног рада. То су влакна по пријему импулса који обезбеђују механизам контракције мишића. Укратко, структура мишића може бити представљена као систем који се састоји од саромерових молекула и миофибрила. Важно је разумети то мишићна влакна формиране од вишеструких подјединица миофибрила, постављених уздужно у односу једна на другу. Сада је потребно посебно размотрити саркомере и филаменте. Зато што играју важну улогу у моторним процесима.
Ad
Сарцомереси и филаменти
Сарцомереси су сегменти влакана који су одвојени такозваним З-плочама које садрже бета-актинин. Из сваке плоче се протежу актински филаменти, а празнине се пуне дебелим аналозима миозина. Елементи актина, заузврат, изгледају као низови перли, уплетених у двоструку спиралу. У овој структури, свака зрна је молекул актина, а молекули тропонина се налазе у подручјима са депресијама у спиралама. Свака од ових структурних јединица формира механизам контракције и релаксације мишићних влакана, који се међусобно повезују. Има кључну улогу у побуђивању влакана ћелијске мембране. Садржи трансверзалне епрувете за инвагинацију које активирају функцију саркоплазматског ретикулума - то ће бити стимулативни ефекат за мишићно ткиво.
Ad
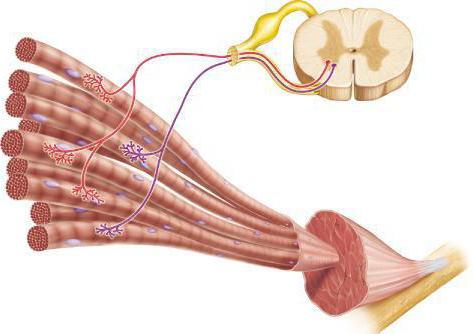
Мотор унит
Сада се исплати удаљити од дубоке структуре мишића и размотрити моторну јединицу у општој конфигурацији скелетног мишића. Ово ће бити скуп мишићних влакана инервираних процесима мотонеурона. Рад мишићног ткива, без обзира на природу дјеловања, осигурат ће се влакнима укљученим у једну моторну јединицу. То јест, када је моторни неурон узбуђен, механизам мишићних контракција се покреће унутар истог комплекса са инервираним процесима. Таква подела на моторне неуроне омогућава намерно коришћење специфичних мишића, без непотребно узбудљивих суседних моторних јединица. У ствари, читава мишићна група једног организма је подељена на сегменте мотонеурона, који се могу ујединити у раду на контракцији или релаксацији, и могу деловати различито или наизменично. Главно је да су независни једни од других и да раде само са сигналима своје групе влакана.
Ad
Молекуларни механизми мишићног рада
У складу са молекуларном концепцијом клизних нити, рад мишићне групе, а посебно његово смањење, остварује се током клизног дејства миозина и актина. Реализован је комплексан механизам интеракције ових нити, у којем се може разликовати неколико процеса:
- Централни део миозин филамента је повезан са сноповима актина.
- Постигнути контакт актина са миозином промовише конформациони покрет молекула потоњег. Главе улазе у фазу активности и развијају се. На тај начин, молекуларни механизми контракције мишића се изводе на позадини реструктурирања нити активних елемената у односу на сваки други.
- Затим долази до међусобног неслагања између миозина и актина, након чега слиједи обнова главе.
Читав циклус се изводи неколико пута, због чега се поменути праменови померају, а З-сегменти саркомере се спајају и скраћују.
Физиолошке особине рада мишића
Међу главним физиолошким својствима мишићног рада, постоји контрактилност и ексцитабилност. Ове особине су, пак, проузроковане проводљивошћу влакана, пластичности и својством аутоматике. Што се тиче проводљивости, она осигурава ширење процеса ексцитабилности између миоцита на нексусу - то су посебни електрично водљиви кругови одговорни за провођење импулса контракције мишића. Међутим, након контракције или релаксације, врши се и рад влакана.
Пластичност је одговорна за њихово мирно стање у одређеном облику, што одређује одржавање сталног тона у којем се тренутно налази механизам контракције мишића. Физиологија пластичности може се манифестовати иу облику очувања скраћеног стања влакана, тако иу облику истезања. Занимљиво је и аутоматизација имовине. Одређује способност мишића да уђу у радну фазу без повезивања нервног система. То јест, миоцити независно производе ритмички понављајуће импулсе за одређена дејства влакана.
Ad
Биохемијски механизми мишићног рада
Цела група хемијских елемената је укључена у рад мишића, укључујући калцијум и контрактилне протеине као што су тропонин и тропомиозин. На основу овог снабдевања енергијом, изводе се физиолошки процеси о којима се горе расправљало. Извор ових елемената је аденозин трифосфат киселина (АТП), и његова хидролиза. У овом случају, залиха АТП у мишићима је у стању да обезбеди контракцију мишића само за делић секунде. Упркос томе, влакна могу да реагују на нервне импулсе у континуираном режиму.
Чињеница је да су биохемијски механизми мишићне контракције и релаксације са АТП подршком повезани са процесом развоја резерве макроерга у облику креатин фосфата. Обим ове резерве је неколико пута већи од АТП залиха и истовремено доприноси његовој производњи. Поред АТП, гликоген може да делује и као извор енергије за мишиће. Иначе, мишићна влакна чине око 75% укупне количине ове супстанце у организму.
Коњугација ексцитаторних и контрактилних процеса
У мировању, влакна влакана не ступају у међусобну интеракцију клизањем, јер су центри лигамената затворени молекулима тропомиозина. Узбуда се може десити тек након електромеханичког повезивања. Овај процес је подељен у неколико фаза:
Ad
- Када се на мембрани миофибрила активира неуромускуларна синапса, формира се такозвани постсинаптички потенцијал који акумулира енергију за акцију.
- Стимулирајући импулс због система цеви дивергира дуж мембране и активира ретикулум. Овај процес у коначници доприноси уклањању баријера из мембранских канала, дуж којих се ослобађају јони везани за тропонин.
- Протеин тропонина, са своје стране, отвара центре актин лигамената, након чега механизам контракција мишића постаје могућ, али за почетак то ће такође захтевати одговарајући импулс.
- Употреба отворених центара ће почети у тренутку када се споје главе миозина према горе описаном моделу.
Комплетан циклус ових операција одвија се у просјеку у року од 15 мс. Период од почетне тачке ексцитације влакана до потпуне редукције назива се латентна.
Процес опуштања скелетних мишића
Када су мишићи опуштени, одвија се обрнути трансфер Ца ++ јона са везаним ретикулумом и калцијумским каналима. У процесу ослобађања јона из цитоплазме смањује се број центара лигамента, што доводи до одвајања актиних и миозинских филамената. Другим речима, механизми контракције мишића и релаксације повезују исте функционалне елементе, али раде са њима на различите начине. Након релаксације, може се јавити процес контрактуре, током којег се посматра стална контракција мишићних влакана. Ово стање може трајати до сљедећег дјеловања иритирајућег импулса. Постоји и контрактура кратког дејства, предуслов за то је контракција тетаником у условима акумулације јона са великим запреминама.

Смањење фазе
Када је мускулатура изазвана иритирајућим импулсом суперграничне снаге, јавља се једна контракција у којој се могу разликовати 3 фазе:
- Већ поменути период редукције је латентног типа, током којег влакна акумулирају енергију за обављање накнадних акција. У овом тренутку, одвијају се електромеханички процеси спајања и отварају се центри лигамента. У овој фази се припрема механизам контракције мишићног влакна, који се активира након ширења одговарајућег импулса.
- Фаза скраћивања траје у просјеку 50 мс.
- Фаза релаксације такође траје око 50 мс.
Редукција мишића
Рад са једном контракцијом је разматран као пример "чисте" механике мишићних влакана. Међутим, у природним условима такав рад се не обавља, јер су влакна у сталном одговору на сигнале моторних живаца. Друга ствар је да, у зависности од природе овог одговора, рад се може десити у следећим режимима:
- Скраћенице се јављају на нижој фреквенцији пулса. Ако се електрични импулс шири након завршетка релаксације, следи низ појединачних чинова контракције.
- Висока фреквенција импулсних сигнала може се подударати са фазом опуштања претходног циклуса. У овом случају, амплитуда у којој је радио механизам контракције мишића би се сабрао, што би омогућило дугорочну контракцију са непотпуним актом релаксације.
- У условима повећања учесталости импулса, нови сигнали ће деловати током периода скраћивања, што ће изазвати продужену контракцију која неће бити прекинута релаксацијом.
Оптимална и максимална фреквенција
Амплитуде контракција су одређене фреквенцијом импулса који иритирају мишићна влакна. У овом систему интеракције сигнала и одговора може се разликовати оптимум и фреквенција фреквенције. Прва је фреквенција, која ће се у тренутку акције надовезати на фазу повећане ексцитабилности. У овом режиму може се активирати велики механизам контракције мишића. Заузврат, пессимум одређује вишу фреквенцију, чији пулс пада на фазу рефракторности. Сходно томе, у овом случају амплитуда се смањује.
Врсте рада скелетних мишића
Мишићна влакна могу радити динамички, статички и динамички инфериорно. Стандардни динамички рад се превазилази - то јест, мишић у тренутку контракције помера објекте или његове саставне делове у простору. Статички ефекат мишића се на неки начин ослобађа стреса, јер у овом случају није предвиђена никаква промена у његовом стању. Динамички-инфериорни механизам контракције скелетних мишића активира се када влакна функционишу под напетошћу. Потреба за паралелним истезањем може бити и због чињенице да рад влакана подразумева извођење операција са спољним телима.
У закључку
Процеси организовања мишићне акције повезују различите функционалне елементе и системе. Рад обухвата комплексан скуп учесника, од којих сваки извршава свој задатак. Може се видети да у процесу активирања механизма контракције мишића раде и индиректни функционални блокови. На пример, ово се односи на процесе генерисања енергетског потенцијала за обављање посла или система блокирања центара лигамената кроз које се јавља веза миозина и актина.
Главно оптерећење пада директно на влакна која обављају одређене радње на командама моторних јединица. Штавише, природа обављања одређеног посла може бити различита. На њега ће утицати параметри вођеног импулса, као и тренутно стање мишића.