Шта је хроматин? Функције хроматина
Генетски материјал еукариотских организама има веома сложену организацију. ДНА молекуле, који се налазе у ћелијском језгру, део су посебне мултикомпонентне супстанце - хроматина.
Дефиниција концепта
Хроматин је материјал ћелијског језгра који садржи наследну информацију, која је комплексан функционални ДНК комплекс са структурним протеинима и другим елементима који обезбеђују паковање, складиштење и реализацију кариотског генома. У поједностављеном тумачењу, ово је супстанца која чини хромозоме. Термин потиче од грчког "хрома" - боје, боје.
Овај концепт је Флеминг увео још давне 1880. године, али још увијек постоје расправе о томе шта је кроматин са становишта његове биокемијске композиције. Несигурност се односи на мали део компоненти које нису укључене у структурирање генетских молекула (неки ензими и рибонуклеинске киселине).
Ad
На електронској фотографији интерфазног језгра, хроматин се визуализује као бројне мрље тамне материје, које могу бити мале и расуте или комбиноване у велике густе накупине.
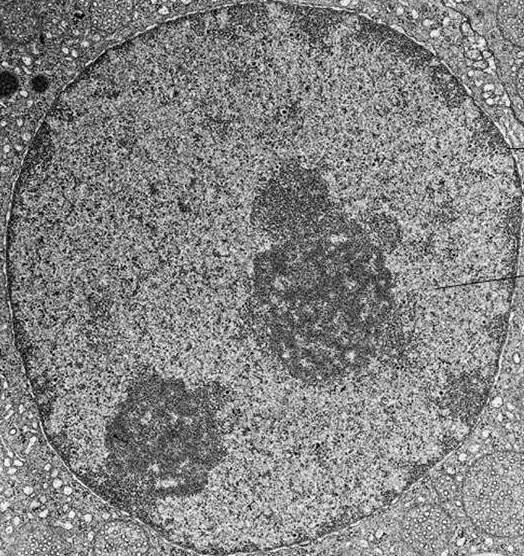
Кондензација хроматина током деобе ћелија доводи до формирања хромозома, који су видљиви чак иу конвенционалном светлосном микроскопу.
Структурне и функционалне компоненте хроматина
Да би се утврдило шта је хроматин на биохемијском нивоу, научници су ову супстанцу извадили из ћелија, пренели у раствор и проучавали састав и структуру компонената у овом облику. Коришћене су и хемијске и физичке методе, укључујући и технологије електронске микроскопије. Показало се да је хемијски састав хроматина за 40% представљен дугим молекулима ДНК и скоро 60% различитим протеинима. Потоњи су подељени у две групе: хистони и не-хистони.
Ad
Хистони су велика породица језгра нуклеарних протеина који се снажно везују за ДНК, формирајући структурални скелет хроматина. Њихов број је приближно једнак проценту генетских молекула.
Остатак (до 20%) протеинске фракције пада на протеине који вежу ДНК и просторно модификујуће протеине, као и ензиме укључене у процес читања и копирања генетичке информације.
Поред главних елемената, у саставу кроматинских рибонуклеинских киселина (РНА), гликопротеини, угљени хидрати и липиди се налазе у малој количини, али је питање њихове повезаности са комплексом за паковање ДНК још увек отворено.
Хистони и нуклеосоми
Молекуларна тежина хистона варира од 11 до 21 кДа. Велики број основних аминокиселинских лизина и аргининских остатака даје овим протеинима позитиван набој, доприносећи формирању ионских веза са супротно набијеним фосфатним групама двоструке спирале ДНК.
Постоји 5 типова хистона: Х2А, Х2Б, Х3, Х4 и Х1. Прва четири типа су укључена у формирање главне структурне јединице хроматина - нуклеозома, који се састоји од језгра (протеинска језгра) и ДНК омотане око ње.
Нуклеосомско језгро је представљено октомерним комплексом осам молекула хистона, који укључује тетрамер Х3-Х4 и димер Х2А-Х2Б. Област ДНК дужине од око 146 нуклеотидних парова је намотана на површину протеинске честице, формирајући 1,75 завојнице, и прелази у линкерску секвенцу (приближно 60 бп) која повезује нуклеосоме један са другим. Х1 молекула се везује за линкер ДНК, штитећи је од дејства нуклеаза.
Ad
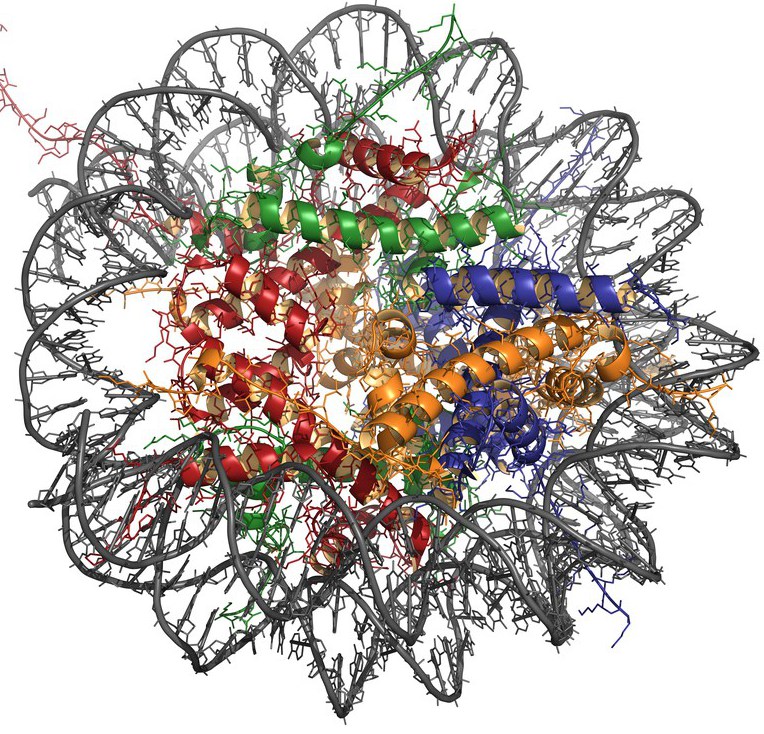
Хистони се могу подвргнути различитим модификацијама, као што су ацетилација, метилација, фосфорилација, АДП-рибозилација и интеракција са убиквитин протеином. Ови процеси утичу на просторну конфигурацију и густину ДНК паковања.
Не-хистонски протеини
Постоји неколико стотина различитих не-хистонских протеина са различитим својствима и функцијама. Њихова молекуларна тежина варира од 5 до 200 кДа. Посебна група се састоји од специфичних протеина, од којих је сваки комплементаран специфичној регији ДНК. Ова група обухвата 2 породице:
- "прстима цинка" - препознају фрагменте од 5 нуклеотидних парова;
- хомодимери су карактерисани структуром хеликса и завојнице у фрагменту који је повезан са ДНК.
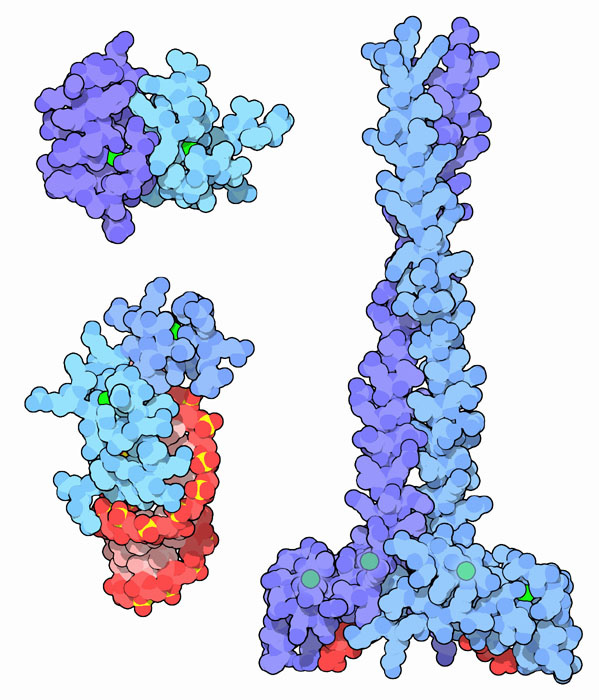
Најбоље су проучавани такозвани протеини високе мобилности (ХГМ протеини), који су трајно повезани са хроматином. Ово име породице примљено је због велике брзине кретања протеинских молекула у електрофоретском гелу. Ова група заузима већину не-хистонске фракције и укључује четири главна типа протеина ХГМ: ХГМ-1, ХГМ-14, ХГМ-17 и ХМО-2. Они обављају структурне и регулаторне функције.
Нехистонски протеини такође укључују ензиме који обезбеђују транскрипцију (синтезу РНК поруке), репликацију (дуплирање ДНК) и поправку (поправку оштећења у генетском молекулу).
Нивои компресије ДНК
Специфичност структуре хроматина је таква да омогућује да се ДНК ланци укупне дужине веће од једног метра уклопе у језгро пречника од око 10 микрона. То је могуће захваљујући вишестепеном систему паковања генетских молекула. Општа шема збијања обухвата пет нивоа:
Ad
- нуклеозомални филамент пречника 10–11 нм;
- фибрил 25–30 нм;
- домени петљи (300 нм);
- Влакна дебљине 700 нм;
- хромозоми (1200 нм).
Овај облик организације обезбеђује смањење дужине оригиналног ДНК молекула за 10 хиљада пута.
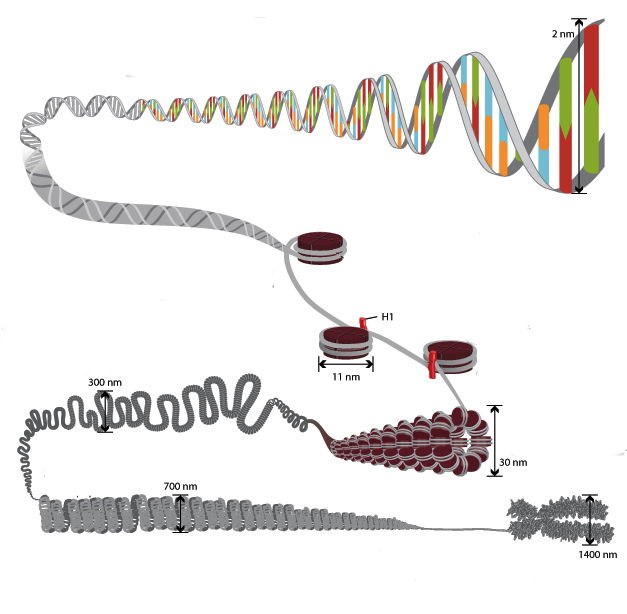
Нит од 11 нм је формиран од стране низа нуклеосома повезаних везним ДНК регионима. На електронским микрографама таква структура личи на перле нанете на линију за риболов. Нуклеосомални филамент се пресавија у калем као соленоид, формирајући фибрил дебљине 30 нм. Хистон Х1 је укључен у његово формирање.
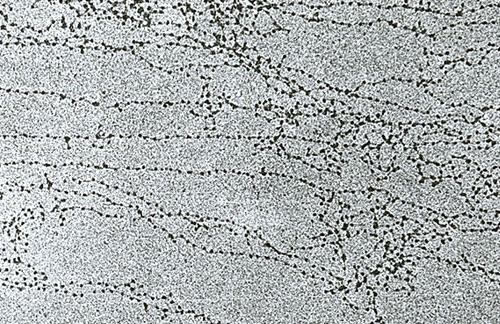
Соленоидна фибрил се савија у петље (тзв. Домене), које су фиксиране на потпорну интрануклеарну матрицу. Сваки домен садржи од 30 до 100 хиљада пари нуклеотида. Овај ниво сабијања је карактеристичан за интерфазни хроматин.
Структура дебљине 700 нм формирана је спиралом фибрила домене и назива се хроматид. Са друге стране, две хроматиде формирају пети ниво ДНК организације - хромозом пречника 1400 нм, који постаје видљив у фази митозе или мејозе.
Ad
Дакле, хроматин и хромозом су облици паковања генетског материјала који зависе од животног циклуса ћелије.
Хромозоми
Хромозом се састоји од две сестринске кроматиде идентичне једна другој, од којих је свака формирана једним суперкованим ДНК молекулом. Половине су повезане специјалним фибриларним телом, названим центромер. У исто време, ова структура је сужавање које раздваја сваку хроматиду на рамена.
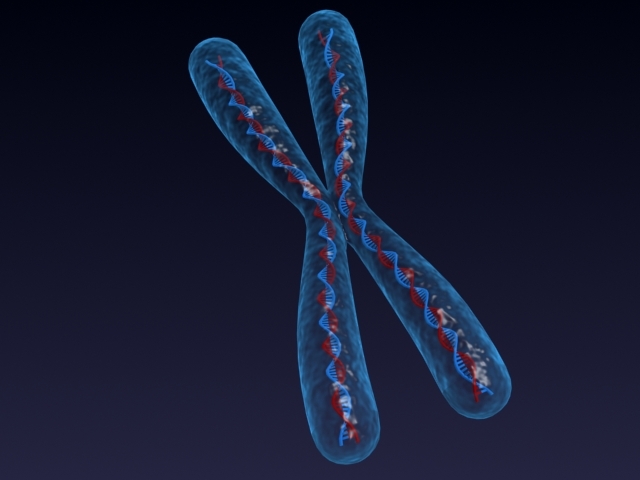
За разлику од хроматина, који је структурни материјал, хромозом је дискретна функционална јединица, коју карактеришу не само структура и састав, већ и јединствени генетски скуп, као и одређена улога у примени механизама наслеђа и варијабилности на ћелијском нивоу.
Еуцхроматин и хетероцхроматин
Хроматин у језгру постоји у два облика: мање спирализован (еукроматин) и компактнији (хетерохроматин). Први облик одговара транскрипцијски активним деловима ДНК и стога није тако чврсто структуриран. Хетерокроматин је подељен на опционо (може да иде од активног до густог неактивног облика у зависности од стадијума животног циклуса ћелије и потребе за применом одређених гена) и конститутивних (стално кондензованих). Током митотске или меиотичке поделе, сви хроматини су неактивни.
Конститутивни хетерохроматин је нађен у близини центромера иу терминалним регионима хромозома. Резултати електронске микроскопије показују да такав хроматин задржава висок степен кондензације не само у фази ћелијске деобе, већ и током интерфазе.
Биолошка улога хроматина
Главна функција хроматина је густо пакована са великом количином генетског материјала. Међутим, једноставно стављање ДНК у језгро није довољно да би ћелија функционисала. Неопходно је да ови молекули исправно "раде", тј. Могу да пренесу информације садржане у њима преко система ДНК-РНК-протеина. Поред тога, ћелија треба да дистрибуира генетски материјал током поделе.
Уређај за хроматин потпуно испуњава ове циљеве. Протеински део садржи све неопходне ензиме, а особитости структуре омогућавају интеракцију са одређеним деловима ДНК. Стога је друга важна функција хроматина да осигура све процесе повезане са имплементацијом нуклеарног генома.