Двострука оплодња у биљкама
Двострука оплодња биљака је од великог биолошког значаја. Навашин га је открио 1898. године. Затим ћемо детаљније размотрити како се двострука оплодња догађа у биљкама. 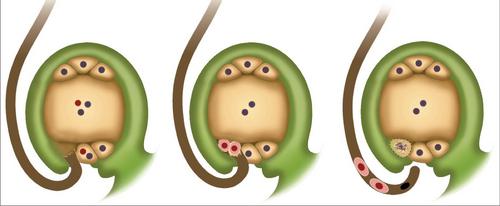
Биолошки значај
Процес двоструке оплодње промовише активан развој хранљивог ткива. С тим у вези, овула не чува материјал за будућност. То, пак, објашњава његов брзи развој.
Шема двоструке оплодње
Укратко, феномен се може описати на следећи начин. Дупла оплодња код ангиосперма састоји се у продирању две сперме у јајник. Један се стапа са јајетом. То доприноси почетку развоја диплоидног ембриона. Друга сперма се спаја са централном ћелијом. Као резултат, формира се триплоидни елемент. Из ове ћелије се појављује ендосперм. То је храњиви материјал за развој ембриона.
Ad
Развој поленске цеви
Двострука оплодња ангиосперми почиње након формирања високо редуковане хаплоидне генерације. Представљена је гаметофитима. Двострука оплодња цветних биљака поспешује клијање полена. Почиње са бубрењем зрна и накнадним формирањем поленске цеви. Она пролази кроз спородерм у својој најтањи дио. Зове се отвор бленде. Од врха поленске цеви емитују се специфичне супстанце. Они омекшавају ткива колоне и стигму. Због тога, они укључују поленску цев. Како се развија и расте, и сперма и нуклеус из вегетативне ћелије прелазе у њу. У великој већини случајева, поленска цев продире у нукелус (мегаспорангиа) кроз микропил јајне ћелије. Ово се ретко ради на други начин. Након продирања у заметну врећицу, поленска се цијев прекида. Као резултат, сав његов садржај се улива унутра. Дупла оплодња цветних биљака наставља се формирањем диплоидног зигота. Ово доприноси првом сперму. Други елемент је повезан са секундарним језгром, који се налази у централном делу ембрионске вреће. Формирана триплоидна језгра се касније трансформише у ендосперм. 
Формирање ћелија: опште информације
Процес двоструке оплодње цветних биљака спроводи се посебним ћелијама. Њихова формација се одвија у две фазе. Прва фаза се зове спорогенеза, друга је хематогенеза. У случају формирања мушких ћелија, ове фазе се називају микроспорогенеза и микрокематогенеза. У формирању женских гениталних елемената, префикс се мења у "мега" (или "макро"). Спорогенеза се заснива на мејози. То је процес формирања хаплоидних елемената. Меиози, као и међу представницима фауне, претходи умножавање ћелија кроз митотске поделе.
Ad
Производња сперме
Примарно формирање мушких гениталних елемената врши се у посебном антерном ткиву. То се зове арцхеспариал. У њему, као резултат митозе, долази до формирања бројних елемената, ћелија мајчиног полена. Затим улазе у мејозу. Услед две мејотске поделе формирају се 4 хаплоидне микроспоре. Неко време леже у близини, формирајући тетраде. Након тога, они се распадају у зрнца полена - појединачне микроспоре. Сваки од формираних елемената почиње да буде прекривен са двије љуске: вањском (екине) и унутрашњом (интин). Онда почиње следећа фаза - микрогаметогенеза. Она се, заузврат, састоји од две митотске узастопне поделе. Након прве две ћелије настају: генеративне и вегетативне. Након тога, први пролази другу подјелу. Као резултат, формирају се две мушке ћелије - ћелије сперме. 
Макропорогенеза и мегаспорогенеза
У ткивима овула, један или више аркспоријских елемената почињу да се раздвајају. Почињу брзо расти. Због ове активности, оне постају значајно веће од осталих ћелија које их окружују у овулама. Сваки елемент арпспоре подвргава се подели митозом један, два или више пута. У неким случајевима, ћелија може одмах да се претвори у мајку. Мејоза се јавља у њему. Као резултат, формирају се 4 хаплоидне ћелије. По правилу, највећи се почиње развијати, претварајући се у заметну врећицу. Три остају постепено дегенерисане. У овој фази, макроспорогенеза је завршена, почиње макро хематогенеза. Током ње се јављају митотске поделе (већина ангиосперма има три). Цитокинеза не прати митосе. Као резултат три поделе, формира се ембрионска врећа са осам језгара. Затим се изолују у независне ћелије. Ови елементи се дистрибуирају на одређени начин помоћу заметне врећице. Једна од изолованих ћелија, која је, у ствари, јајна ћелија, заједно са још два синергида, одвија се на микропилу, у који се одвија пенетрација сперме. У овом процесу, синергисти играју веома значајну улогу. Они садрже ензиме који доприносе растварању мембрана на поленским цевима. На супротној страни клица, остале три ћелије су лоциране. Зову се антиподи. Уз помоћ ових елемената, хранљиве материје се преносе из овула у заметну кесицу. Преостале две ћелије се налазе у централном делу. Често се спајају. Као резултат њихове повезаности формира се диплоидна централна ћелија. Након што се деси двострука оплодња, а сперматозоиди продру у јајник, један од њих ће се, као што је већ речено, спојити са ћелијом јајета. 
Садржи цев од полена
Дупла оплодња је праћена интеракцијом са спорофитним ткивима. То је сасвим специфично. Овај процес је регулисан активношћу хемијских једињења. Утврђено је да ако се полен испере дестилована вода изгубиће способност да клија. Ако се добијени раствор концентрише и затим обради, поново постаје комплетан. Развој поленске цеви након клијања контролише ткиво пиштоља. На пример, у памуку његов раст до јаја траје око 12-18 сати. Међутим, већ након 6 сати сасвим је могуће утврдити на коју ће јајовцу поленска цијев ићи. То је разумљиво јер у њему почиње уништавање синергије. Тренутно није утврђено како биљка може усмерити развој цијеви у правом смјеру и како синергист учи о апроксимацији. 
"Забрана" на самоубиство
Често се примећује у цветним биљкама. Овај феномен има своје карактеристике. „Забрана“ самооплодње се манифестује у чињеници да спорофит „идентификује“ сопствени мушки хематофит и не дозвољава му да учествује у оплодњи. Истовремено, у неким случајевима, полен пиштоља не проклија свој пелуд. Међутим, по правилу, раст цеви и даље почиње, али се касније зауставља. Као резултат, пелуд не допире до јаја и као резултат тога, не долази до двоструке оплодње. Дарвин је приметио овај феномен. Дакле, нашао се у јаглацу спринг фловерс два облика. Неки од њих су били дуге стабљике са кратким прашницима. Други су кратки. У њима су трачасти филаменти били дугачки. Кратко стабљике се разликују у великом полену (двоструко више него у другим). У исто време, ћелије у папилама стигме су мале. Ове знаке контролише група уско тканих гена. 
Рецептори
Дупла оплодња је ефикасна када се полен преноси из једног облика у други. За препознавање сопствених елемената одговорни су посебни молекули рецептора. То су комплексна једињења угљених хидрата са протеинима. Утврђено је да се облици дивљег купуса, који не производе ове рецепторске молекуле у ткивима стигме, могу самоплинирати. За нормалне биљке, једињења угљикохидратних протеина појављују се дан прије отварања цвијета. Ако отворите пупољак и обрадите га сопственим поленом два дана пре његовог цветања, онда ће доћи до двоструке оплодње. Ако се то ради дан прије отварања, неће бити.
Ad
Аллелес
Важно је напоменути да се у неким случајевима "само-некомпатибилност" полена у биљкама успоставља низом вишеструких елемената једног гена. Овај феномен је сличан некомпатибилности трансплантације ткива код животиња. Такви алели су означени словом С. Број становника у овим елементима може досећи десетине или чак стотине. На пример, ако је генотип фабрике за производњу јаја с1с2, а с2с3 за производњу полена током унакрсног опрашивања, клијање ће бити забележено само у 50% честица прашине. То ће бити они који носе алел с3. Ако постоји неколико десетина елемената, већина полена ће нормално клијати током унакрсног опрашивања, док је самооплодња потпуно спречена. 
У закључку
За разлику од гимносперми, које карактерише развој довољно снажног хаплоидног ендосперма, без обзира на оплодњу, код ангиосперма, ткиво се формира само у овом случају. С обзиром на огроман број генерација, тиме је остварена значајна уштеда енергије. Повећање степена плоидности ендосперма, очигледно, доприноси бржем расту ткива у поређењу са диплоидним слојевима спорофита.